肝细胞
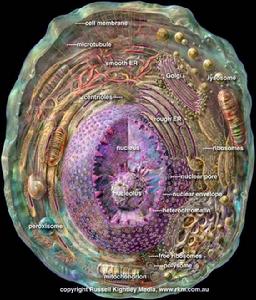
肝脏是由肝细胞组成,肝细胞极小,肉眼看不到,必须通过显微镜才能看到。人肝约有25亿个肝细胞,50个肝细胞组成一个肝小叶,因此人肝的肝小叶总数约有50万个。肝细胞为多角形,直径约为20-30/加(微米),有6-8个面,不同的生理条件下大小有差异,如饥饿时肝细胞体积变大。每个肝细胞表面可分为窦状隙面、肝细胞面和胆小管面三种。肝细胞里面含有许许多多复杂的细微结构:如肝细胞核、肝细胞质、线粒体、内质网、溶酶体、高尔基氏体、微粒体及饮液泡等组成。
目录 |
简介
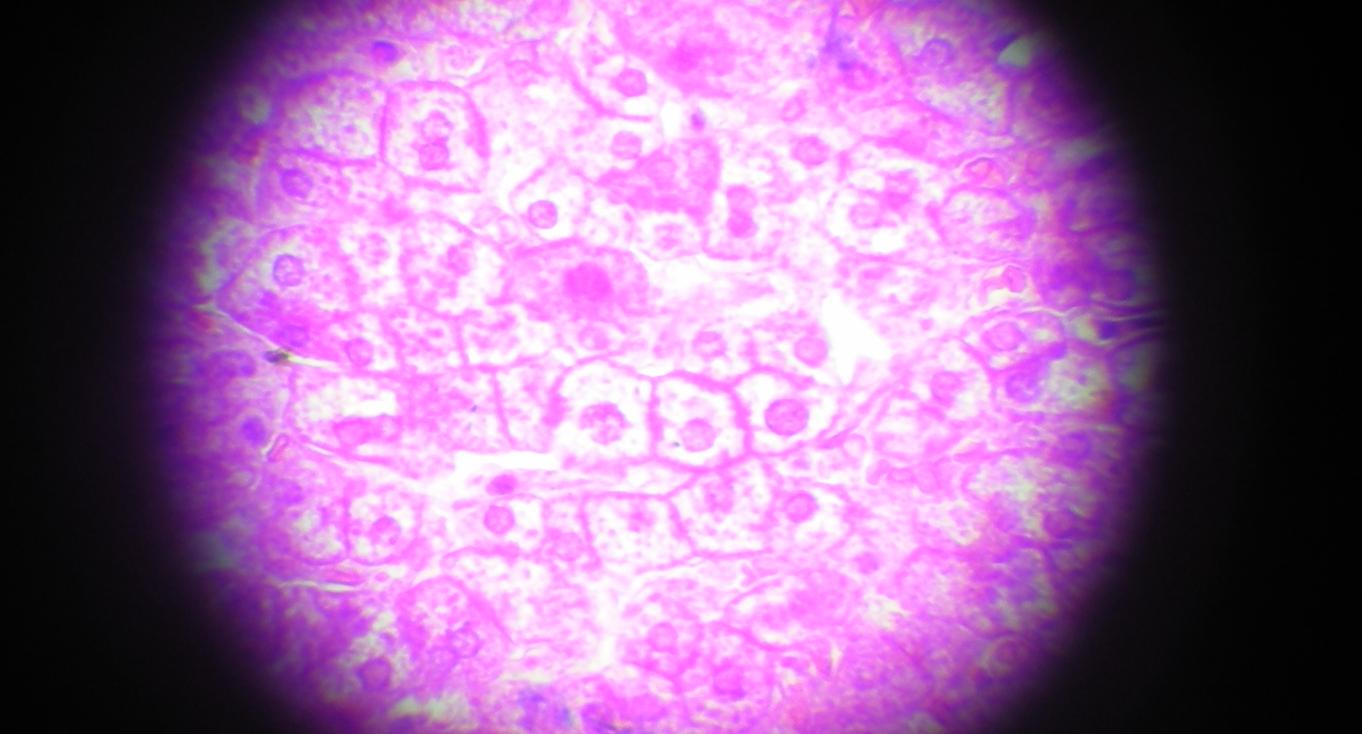
肝细胞hepatocyte,hepatic cells,liver cells
LM(光镜):多面体形;核大而圆,居中,常染色质丰富,部分有双核或多倍体核;胞质嗜酸性,含弥散分布的嗜碱性团块
EM(电镜):
(1)有三种功能面→血窦面
→胆小管面
(2)细胞器发达
粗面内质网:合成白蛋白、纤维蛋白原、凝血酶原、脂蛋白和补体等血浆蛋白
滑面内质网:参与生物转化和代谢,如胆汁合成、脂类代谢、糖代谢、激素代谢和有机异物的转化
线粒体、溶酶体和过氧化物酶体丰富
(3)含糖原、脂滴、色素等内涵物
肝细胞核
肝细胞核主要由去氧核糖核酸(DNA)和组蛋白等组成。去氧核糖核酸是遗传的物质基础,它有复制遗传信息的功能。患肝炎时,肝炎病毒侵入肝细胞核内,病毒基因可以与肝细胞核中去氧核糖核酸相结合(整合)。一旦整合,HBsAg即难以清除,致使HBsAg长期携带。此外,去氧核糖核酸还可能以自己为模板合成信使核糖核酸(mRNA),从而控制细胞质中各种相应蛋白质的合成。肝细胞核如果明显受损,就意味着整个肝细胞崩解毁灭。
线粒体
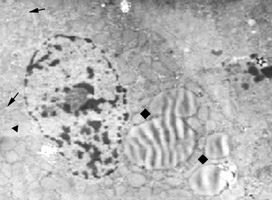
肝细胞的线粒体很多,每个细胞大约有1000个左右,遍布于胞质内。肝小叶不同部位肝细胞内线粒体的大小和形态不完全一致,在正常生理条件下,多为圆形和卵圆形,直径0.4-0.8μm。线粒体的共同基本形态结构特征是外被双层界膜--外界膜和内界膜,内界膜向线粒体内部伸展转折,形成许多嵴。内界膜将线粒体分隔为内、外两室,外室介于内、外界膜之间,内室则围于内界膜之间,其中充满基质。
在线粒体嵴的表面和内界膜的基质面上附有密集排列的、火柴头状的亚单位,称为基粒。其粒由约10nm的球形头部与宽约3.5nm、长约5nm的短柄构成。头部伸入基质,相当于ATP合成酶所在处,是氧化磷酸化最终合成ATP酶的一部分。柄的一端与嵴的界膜相连,是一联接蛋白,脂类和氨基酸在线粒体内被氧化成水和二氧化碳,所释放的能量则通过这种连接蛋白转给ADP生成ATP。线粒体的亚单位只有在特殊处理的标本中,如将线粒体由细胞匀浆中分离出来,并用低渗法破坏其界膜,使其嵴暴露,再用磷钨酸复染,方可看到。而普通超薄切片则见不到这种亚单位,这是因为锇酸固定时,球形头部被解聚所致。
线粒体内外界膜的通透性和化学组成互不相同。外界膜对大多数分子量小于10000的低分子溶质而言,均可自由通过,而内界膜则仅允许不带电荷、相对分子量小于150的小分子如水、O2、CO2、尿素及甘油等通过,葡萄糖、K+、Na+、Cl-等均不能通过内膜。线粒体基质内常见一些小的电子致密颗粒,称为线粒体内颗粒或基质颗粒。基质颗粒内含Ca2+、Mg2+等离子。
线粒体基质内含有蛋白质(包括各种酶类、类脂质成分、DNA、RNA及核蛋白体),除此之外,还有各种单核苷酸和辅酶。由于线粒体膜和基质内含有大量酶类,如含有进行氧化作用的呼吸链的酶体系,氧化磷酸化酶体系、三羧酸循环及脂肪酸氧化的酶体系等,各种代谢物质在线粒体内氧化,并把放出的能量转换成ATP。因此,线粒体像一个能源中心,一个电力发动站,能向细胞不断提供其生命活动所必需的能量,以保证和推动细胞进行各种复杂的生理功能。
由于线粒体还含有自身的基因物质——脱氧核糖核酸,称线粒体脱氧核糖核酸(mt DNA)或称染色体外脱氧核糖核酸其含量约占一个细胞全部DNA的2%。此外,线粒体内还含有DNA聚合酶,KNA聚合酶,氨基酸活化酶,tRNA及mRNA。因而能自我复制和合成蛋白质,自行分裂、繁殖和增生。线粒体是细胞内最为敏感的细胞器之一。在许多病理情况下,线粒体常常首先发生各种形态改变,最常见的有线粒体肿胀,线粒体增生、肥大以及线粒体内包含物的产生等。
内质网
肝细胞的内质网很丰富,分布广泛。它与高尔基复合体及核膜共同构成一连续的细胞内网状膜系统。按其囊膜表面是否附着核蛋白体,将内质网分为两类:即粗面内质网及光面内质网。

粗面内质网
粗面内质网RER形成池(cistem),在其膜外面附着有核蛋白体,据测定,1mg肝组织中所含内质网的总面积约为11m2,其中约2/3为RER。在肝小叶的不同区带其分布是不相同的。肝小叶周边带的肝细胞内,RER尤为丰富。光面内质网则相反,在小叶中央带及中间带肝细胞内较小叶周边带肝细胞内为多。这种数量上的分布差异与小叶不同区带内肝细胞的功能特性有关。在一个细胞中,RER的主要功能为生成输出蛋白(或称分泌蛋白),如血浆白蛋白、α、β球蛋白、纤维蛋白原、凝血酶原等,均在RER上合成。新合成的蛋白质贯穿内质网膜进到内腔,经由运输小泡运至高尔基复合体,在此加工、浓缩、再经分泌泡从细胞表面释放入肝窦内。RER的发达程度反映着肝细胞的功能状态。各种损伤因子引起的肝细胞损伤,亦可反映于RER,其最常见的改变为RER膜上多聚核蛋白体解聚(disaggregation)及脱粒(degranulation)。解聚是指多聚核蛋白体分散为单体,游离分散在细胞质中,或附在粗面内质网膜上。脱粒则指附着在RER膜上的核蛋白体脱落下来,多以单体形式散在胞质之中。如四氯化碳中毒所致肝细胞损害时,可见RER膜上多聚核蛋白体解聚及脱粒,此时,蛋白质合成也聚降。肝癌时,RER数量与肿瘤细胞的生长率及恶性程度之间存在一种反相关关系,在分化较高、生长缓慢的癌细胞中,RER较发达,反之,在分化低,生长迅速的肝癌细胞中,RER则往往很少,而游离的多聚核蛋白体却十分丰富,以适应癌细胞快速生长的需要。
光面内质网
光面内质网(SER)膜上不附有核蛋白体,由分支的小管组成,并与RER相连系,也可由RER形成。SER在肝细胞中具多种功能,如参与糖原代谢、胆汁分泌、脂类代谢、类固醇激素代谢及解毒等。SER常位于胞浆一侧,与糖原颗粒相伴随,若糖原很丰富时,常可遮盖SER使之分辨不清。在饥饿时,糖原减少,肝细胞内SER则较为明显。由于SER含有葡萄糖-6磷酸酶,加之与糖原结构空间的紧密关系,因而更有利于糖原代谢。
SER与胆汁分泌有关,非结合性胆红素从血液进入肝细胞后,经SER上的葡萄糖醛酸转移酶作用,成为水溶性结合胆红素而利于排泄,胆盐也在SER上合成。此外,SER还参与脂肪代谢。血液内游离脂肪酸进入肝细胞后,在SER上酰化成甘油三酯,经与RER合成的蛋白质结合,形成极低密度脂蛋白进入血窦。
肝脏的解毒功能亦在SER上进行。由于SER内含有混合功能氧化酶系,其中终末氧化酶即细胞色素P-450,对许多有害物质如机体代谢产物、药物、致癌剂、杀虫剂等均可加以代谢,从而或被解除毒性,或被转化为易于排泄的物质。由此不难理解,在慢性药物中毒过程中(如安眠药、巴比妥类中毒)可见到SER膜的增生。SER膜的增生还见于长期用抗组织胺药物,口服抗糖尿病药物和避孕药时。SER膜的增生,一般属细胞的一种适应性反应,是功能升高的表现(即酶的诱导)。但并非任何SER的增生均伴有功能的升高,有时往往表现为一种无效增生。在胆汁淤积时,肝细胞内增生的SER则处于低活性状态。表面抗原阳性的乙型肝炎病人,也出现肝细胞内光面内质网增生,在其小管内形成乙型肝炎表面抗原。此时的肝细胞由于含有增生的光面内质网,在组织切片上模糊如毛玻璃,故称毛玻璃细胞。电镜下,可见光面内质网小管中心呈细丝状的乙型肝炎表面抗原。这在诊断上颇为重要。
核蛋白体

核蛋白体(ribosome)又称核糖核蛋白体或核糖体,因首先被Palade在电镜下发现,故又称palade小体。
核蛋白体可游离于胞浆基质中,称游离核蛋白体,亦可附着在内质网膜上,构成粗面内质网。肝细胞具丰富的游离核蛋白体,由60s和40s(S=Svedberg,沉降系数的单位)的大、小二个亚单位组成,呈颗粒状。大亚单位直径约为23μm,略呈锥体形,内含一中央管,底边扁平,有一窄沟。小亚单位略呈弧形(23nm×12nm),一面外凸,一面凹陷;在镁离子存在时,大小亚单位结合成单核蛋白体,此时,小亚单位的凹面与大亚单位的扁平底面相贴,小亚单位的中间分界线正与大亚单位底面的沟相吻合成隧道。
核蛋白体的主要成分为核蛋白体核糖核酸,它们与蛋白质结合,以核蛋白(RNP)的形式存在。
核蛋白体可以单个存在,即单体,也可以由mRNA细丝将它们串联一起,构成多聚核蛋白体。多聚核蛋白体是合成蛋白质的功能基团,mRNA穿行于大小亚单位之间的隧道中,新合成的肽链便自中央管释放出来。当特定的主链形成后,核蛋白体便从mRNA上离去,并分解成亚单位。需要合成蛋白质时,亚单位再行结合,并进一步组成多聚核蛋白体。因此,在细胞质中,核蛋白体的单体和多聚体总是随着细胞的功能状态处于不断结合和分解的动态变化之中。游离核蛋白体合成的蛋白质主要供肝细胞自身生长、分裂、更新所需。
溶酶体
DeDuve于1955年首次在大鼠肝细胞匀浆超速离心后的各组成分中发现溶酶体的存在,后经电镜观察证实。溶酶体是由单层界膜围成的颗粒,其大小、形态以及内部结构均极不一致。由于所有溶酶体均含有酸性水解酶,故将此酶作为溶酶体的标志酶。溶酶体借助其所含50多种酶消化、分解各种内生性或外源性物质,因此,可将其视为细胞内的消化器官。肝细胞内所含溶酶体较为丰富,根据其是否含有作用底物而分为两种:初级溶酶体(primarylysosome)和次级溶酶体(secondarylysosome)。

初级溶酶体
此类溶酶体仅含水解酶而无底物,由单层界膜包绕,内含电子致密的均质物,常位于近高尔基复合体处。初级溶酶体在粗面内质网上合成,经运输小泡送至高尔基囊泡进行加工、浓缩,再由高尔基扁平囊分泌面末端膨大、分离而形成初级溶酶体。溶酶体所含水解酶能消化各类大分子化合物。在正常生理情况下,该种酶处于非激活状态,同时,溶酶体膜的内表面还有一层带电荷的糖蛋白,保护膜不受水解酶的作用。而且,溶酶体膜还具有独特的滤过性质,只允许分子量小的物质通过。这就保证了大分子的水解酶不能自由逸出膜外,从而保护细胞免于自身消化。
次级溶酶体
此类溶酶体内除含有水解酶外,还含有相应的作用底物,以及由此形成的消化产物。由于所含底物的不同和消化程度的差异,构成了次级溶酶体形态的多样性。根据消化底物来源的不同,又可将次级溶酶体分为自生性、异生性和兼性三种。
高尔基氏体
电镜下,高尔基复合体(Golgicomplex)由三种基本成分组成即扁平囊泡、小泡和大泡,多位于细胞核与毛细胆 管间的区域内。

扁平囊泡
扁平囊泡(saccule)由一组弯曲呈蹄铁形的扁平囊泡组成,来源于核膜外层。弯曲的囊泡有两个面(凸面和四面),凸面又称形成面(forming face),或称未成熟面(immatureface),靠近胞核;凹面为分泌面(secreting face),或称成熟面(matureface),面向细胞膜。形成面的囊膜较薄,近似细胞膜。因此,高尔基囊泡可视为内质网膜与细胞膜的中间分化阶段。
小泡
小泡(vesicle)数量较多,与一般胞饮小泡相似,常散布于扁平囊泡的形成面,小泡由高尔基复合体附近的粗面内质网芽生而来,并载有粗面内质网所合成的蛋白质成分,后者被运送到高尔基复合体的囊泡形成面,在此,小泡与形成面的扁平囊泡膜融合,蛋白质乃进入囊泡腔中。
大泡
大泡(vacuole)为扁平囊泡末端局部膨大而成,又称分泌泡或浓缩泡,大泡带着由扁平囊泡所生长的分泌物质(如脂蛋白、胆汁成分)断离扁平囊泡,将其运往窦腔或排向毛细胆管。分泌面细胞膜周微丝、微管系统的存在,是这一功能得以实现的必要前提和保证。小泡的并入及大泡的断离,使高尔基囊膜不断处于新陈代谢的动态变化之中。
常见的高尔基复合体病理改变为肥大或萎缩。高尔基复合体肥大多见于分泌障碍并伴有高尔基大泡内分泌物潴留及淤胆等。胆汁成分潴留于分泌泡中;在营养性或中毒性肝脂肪变时,脂蛋白潴留于分泌囊泡中。高尔基复合体萎缩则常见于核蛋白体的合成功能下降状态,如各种毒性因子造成的肝细胞蛋白合成降低时,即常见粗面内质网脱颗粒和断裂。并伴有高尔基复合体的萎缩或消失。
微体
微体是肝细胞内最小的细胞器,为由单层界膜包绕的圆形或卵圆形小体。在肝细胞内,微体与线粒体的比例近乎1:4。微体基质内含有过氧化氢酶和多种氧化酶,如D-氨基酸氧化酶、L-氨基酸氧化酶及L-2-羟基酸氧化酶等,故微体又称过氧体(peroxisome)。
过氧体来源于粗面内质网,形成迅速,从粗面内质网转运出来大约只需一小时便可完成,在细胞内可存在5天,并在4分钟内通过自噬或自溶过程而解体。亦有人认为,微体尚可合并到溶酶体或衍化成线粒体。从微体的发生及其所含酶的特点看,可视为一种特殊类型的溶酶体。从种系发生史上看,微体可被视为一种古老的氧化产能微器官的遗迹,在细胞生物进化过程中逐渐被线粒体所取代。
微体内含有对长链脂酸进行β氧化降解的酶系,故能参与脂代谢。微体内的过氧化氢酶能降解细胞内的H2O2以防止氧化氢引起细胞中毒。微体内的过氧化氢酶和L-2-羟基酸氧化酶能将NADH再氧化,并通过过氧体的α-甘油磷酸脱氢酶,支持果糖的降解。因此,微体和线粒体是协同参与细胞呼吸的细胞器。
微体与微粒体(microsome)很易混淆,但这是两个完全不同的概念。微体是细胞内固有的细胞器,而微粒体则系缀有大量核糖体的内质网碎片,是组织匀浆超速离心后的产物。在病理情况下,可见有肝细胞内微体数目的增多或减少。微体增多可由甲状腺索引发,因而在甲状腺功能亢进患者的肝细胞内,常见有微体数量的增多;反之,甲状腺功能低下时,肝细胞内微体数目则减少。此外,慢性酒精中毒及肝癌时亦均见有肝细胞内微体数目的增多,其意义尚不清楚。
肝细胞实验
肝细胞:68号切片
低倍镜下找到呈多边形的肝小叶,选择一个肝小叶换高倍镜观察,可见到呈索状排列的肝细胞,呈多边形,有1-2个圆形细胞核,核仁明显,核膜清楚,核内染色质稀疏,染色较浅
观察细胞器与内含物
细胞器与内含物的种类很多,实验课仅观察几种主要的细胞器和内含物在光镜下的形态和位置。各种细胞器和内含物,在一般的HE染色的切片上看不到,须用特殊方法染色显示。
线粒体——示教:3号片
小狗胰脏,Regaud氏液固定,石蜡切片,铁苏木素染色。
线粒体用铁苏木素染色呈黑色,分布于核周围的细胞质中,线粒体在高倍镜下呈粒状、线状或短棒状,或直或曲,轮廓鲜明。
胰脏的分泌细胞呈锥形,核大而圆,位于细胞中央,细胞游离端聚集有许多大而圆的黑色颗粒为分泌颗粒。
高尔基复合体——示教:2号片
在切片上有大小不一的圆形脊神经节细胞,胞核不着色,但能看到淡黄色的核仁。核周围的细胞质中有镀染成黑色的网状或颗粒状物,即光镜下的高尔基复合体。
糖元——示教:6号片
猪肝,冷Carnoy氏液固定,PAS反应法显示糖元,苏木素复染胞核。
在高倍镜下,肝细胞中可见到大量紫红色的糖元颗粒或小块,在很多肝细胞中,糖元偏于细胞一侧,此系制片过程造成,生活状态时分布较均匀。
| ||||||||||||||||